- 首页
- 产品
-
免疫检查点蛋白
细胞因子靶点
全长多次跨膜蛋白
-

免疫组化 (IHC) 抗体
同型对照
-

ELISA
qPCR试剂盒
流式细胞术
ADC 偶联试剂盒
免疫组化(IHC)试剂盒
MPCLIA
-

新冠病毒
水痘-带状疱疹病毒
-

实验仪器设备
-
-
通用型蛋白
药物筛选与分析服务
-

细胞培养产品
Premium(Pre-GMP) & GMP 级别原材料
基因编辑产品
-

残留检测试剂盒 - resDetect
-
审计&申报文件支持
-
-
-

Premium(Pre-GMP) & GMP 级别原材料
基因编辑
-

残留检测 - resDetect
-
-

-

残留检测 - resDetect
-

免疫原性研究
审计&申报文件支持
-
-
-
ADC研发工具
-

免疫原性研究
审计&申报文件支持
-
-
-

基因编辑
mRNA合成
-

抗原含量检测
疫苗生物效价检测
残留检测试剂盒 - resDetect
-

疫苗生物效价检测
-

疫苗生物效价检测
-

-
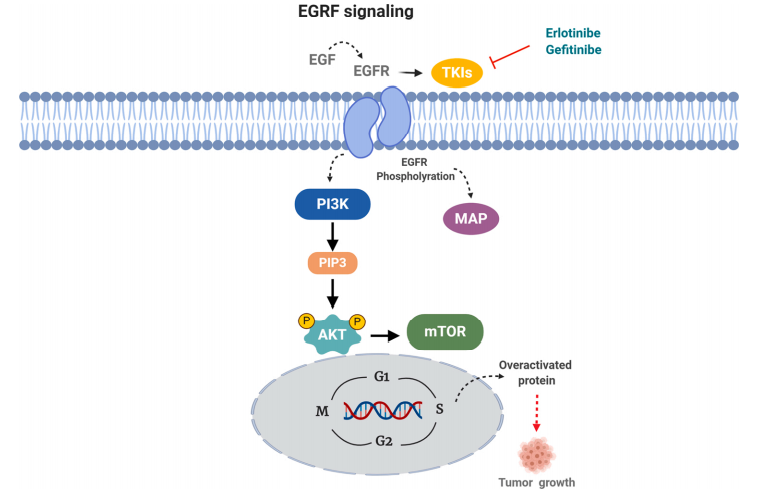
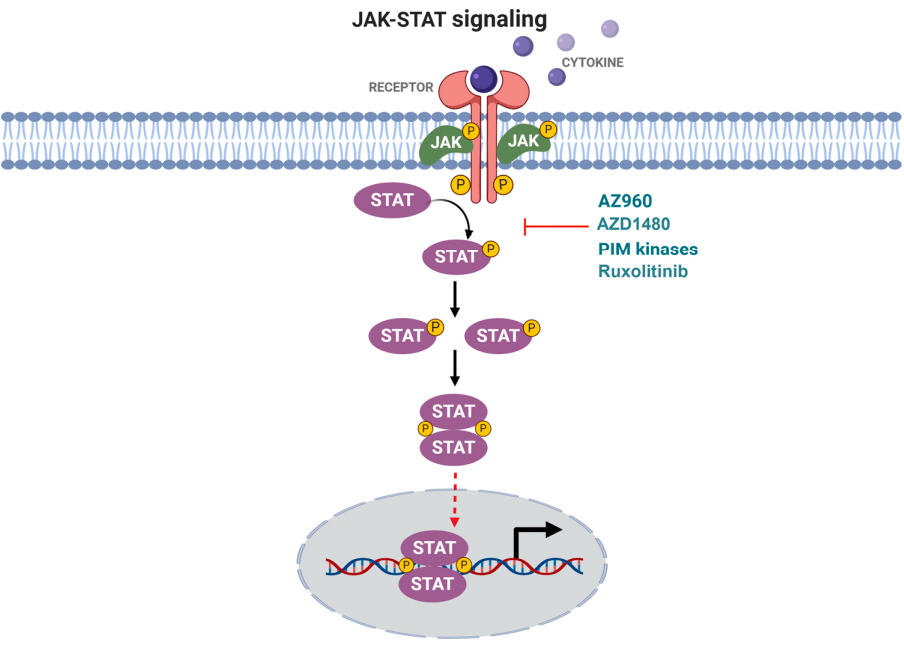
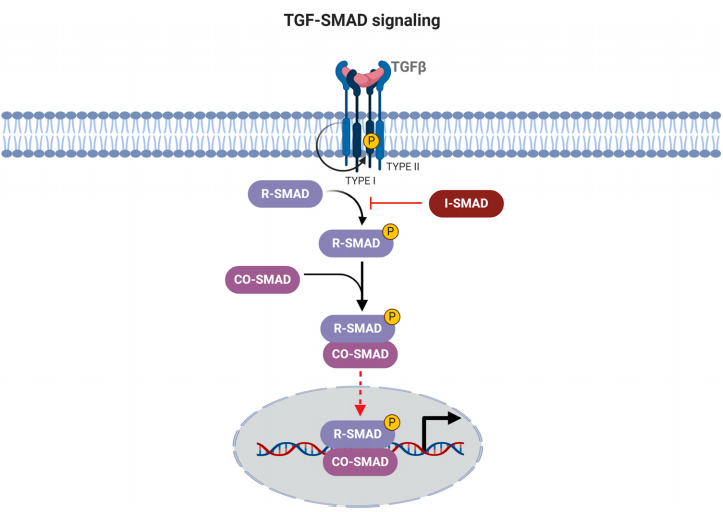
 Star Ribbon预染蛋白Marker蛋白质标记物是生物研究和药物开发的重要组成部分。无论是用于蛋白质电泳还是western blot,我们的预染色蛋白质标记物帮助您快速确定目标蛋白质的分子量或评估转移效率。Fc受体蛋白治疗性抗体的功效取决于Fab片段及其对目标抗原的结合活性,还取决于Fc片段及其与关键Fc受体的相互作用。因此,在抗体工程中候选物必须针对一系列受体进行测试。探索我们的重组Fc受体蛋白质的全面收藏!
Star Ribbon预染蛋白Marker蛋白质标记物是生物研究和药物开发的重要组成部分。无论是用于蛋白质电泳还是western blot,我们的预染色蛋白质标记物帮助您快速确定目标蛋白质的分子量或评估转移效率。Fc受体蛋白治疗性抗体的功效取决于Fab片段及其对目标抗原的结合活性,还取决于Fc片段及其与关键Fc受体的相互作用。因此,在抗体工程中候选物必须针对一系列受体进行测试。探索我们的重组Fc受体蛋白质的全面收藏!
-
- 技术服务
- 质量&平台
-
技术资源
- 会议&合作

- 市场活动

- 公司
























 膜杰作
膜杰作 Star Staining
Star Staining